 |
 |
 |
 |



Prey Abundance, Space Use, Demography, and Foraging Habitat of Northern
Goshawks in Western Washington
|
Here's a link to the PDF version of this Thesis
Thomas David Bloxton, Jr
A thesis submitted in partial fulfillment of the
requirements for the degree of Master of Science
Program Authorized to Offer Degree: College of Forest Resources
TABLE OF CONTENTS
List of Figures Chapter 2: Foraging Behavior and Habitat
Use by Northern Goshawks in
ACKNOWLEDGEMENTSThis research would not have been possible without the cooperation and commitment of many public agencies and private companies and the dedication of their personnel. The Washington State Department of Natural Resources (DNR), Rayonier, Champion Pacific Timberlands, Inc., Port Blakely Tree Farms, Weyerhaeuser, Sustainable Ecosystems Institute, Washington Department of Fish and Wildlife, Olympic National Park, U.S. Forest Service - Pacific Northwest Research Station, Plum Creek Timber Company, International Paper, Crown Pacific, Merrill and Ring Tree Farms, Campbell Group, Olympic Resources Management, Sarvey Wildlife Center, Sardis Wildlife Center, Northwest Raptor Center, and the University of Washington's Olympic Natural Resources Center provided funding, access to lands, and logistical support. The following people were indispensable in my efforts during the course of study: R. Meier, L. Raynes, L. Young, P. Swedeen, J. Eskow, A. Stringer, N. Wilkins, K. Risenhoover, T. McBride, D. Runde, L. Hicks, H. Stabins, F. Silvernail, S. Horton, P. Harrison, E. Kuo-Harrison, D. Hays, S. Desimone, P. Happe, K. Jenkins, S. Hall, S. Gremel, and J. Swingle. I also thank the following people for their diligent efforts and assistance in the field: D. Yonkin, B. Davies, J. Luginbuhl, E. Neatherlin, J. Bradley, B. Griffith, H. Tall, K. Beesley, J. DeLap, C. Wilkerson, D. D'emilio, S. Pollock, J. Bent, D. Pike, and P. Smith. Graduate committee members M. Restani and J. Agee provided invaluable guidance. I especially thank S. Finn for his dedication to the initial phase of this project from 1996-1998 and for his assistance throughout the years that followed. Finally, I wholeheartedly thank J. Marzluff and D. Varland for their unending guidance and support during this valuable learning experience.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Year(s) studied for:
|
|||||
|
Territory
|
Sex
|
Landscape
History |
Space Use
|
Survivorship
|
Reproduction
(by territory) |
| Burnt Mountain |
M*
|
2,3
|
1998
|
1997-1999
|
1996-1999
|
| Burnt Mountain |
F*
|
2,3
|
1998
|
1997-1999
|
-
|
| Dungeness |
M*
|
2
|
19,972,000
|
1997-2000
|
1996-2000
|
| Dungeness |
F*
|
2
|
1997
|
1997-1998
|
-
|
| Raney Creek |
M*
|
3
|
1996-2000
|
1996-2000
|
1996-2000
|
| Raney Creek |
F*
|
3
|
1996-1997
|
1996-1997
|
-
|
| The Hole |
M*
|
3
|
1998-1999
|
1998-2000
|
1996-2000
|
| The Hole |
F*
|
3
|
1998
|
1998-1999
|
-
|
| Lily Creek |
M*
|
3
|
1997,1999-2000
|
1997-2000
|
1997-2000
|
| Lily Creek |
F*
|
3
|
1997
|
1997-1999
|
-
|
| Rushingwater Creek |
M*
|
3
|
1998
|
1998
|
1998-2000
|
| Rushingwater Creek |
F*
|
3
|
1998-1999
|
1997-1999
|
-
|
| Swift Creek |
M*
|
1,3
|
2000
|
2000
|
1998-2000
|
| Swift Creek |
F*
|
1,3
|
2000
|
2000
|
-
|
| Snoqualmie |
M*
|
2,3
|
1999-2000
|
1997-2000
|
1997-2000
|
| Snoqualmie |
F1*
|
2,3
|
-
|
1997-1998
|
-
|
| Snoqualmie |
F2*
|
2,3
|
2000
|
2000
|
-
|
| Big River |
M*
|
3
|
1999
|
1999-2000
|
1999-2000
|
| Big River |
F
|
3
|
-
|
-
|
-
|
| Mt. Zion |
M
|
2
|
-
|
-
|
1997-2000
|
| Mt. Zion |
F*
|
2
|
-
|
1997-2000
|
-
|
| St. Paul's Lookout |
M*
|
3
|
-
|
1997-1999
|
1997-2000
|
| Lake Cavanaugh |
M*
|
3
|
-
|
1999-2000
|
1999-2000
|
| Lake Cavanaugh |
F*
|
3
|
-
|
1999-2000
|
-
|
| Calawah/Sitkum |
M*
|
1,2
|
-
|
1997-1998
|
1996-1998
|
| Calawah/Sitkum |
F
|
1,2
|
-
|
-
|
-
|
| North Fork Solduc |
M*
|
1,2
|
-
|
1997-1998
|
1997-2000
|
| North Fork Solduc |
F*
|
1,2
|
-
|
1997-1998
|
-
|
| Lillian River |
M*
|
1
|
-
|
2000
|
1998-2000
|
| Lillian River |
F*
|
1
|
-
|
2000
|
-
|
| Mueller Creek |
M*
|
1,2
|
-
|
1997-1998
|
1997-1998
|
| Mueller Creek |
F*
|
1,2
|
-
|
1997
|
-
|
| Mowich River |
N/A
|
1,3
|
-
|
-
|
1998-2000
|
| Thunder Lake |
N/A
|
3
|
-
|
-
|
1999-2000
|
| Donkey Creek |
N/A
|
3
|
-
|
-
|
1996-2000
|
| Boundary Creek |
N/A
|
2
|
-
|
-
|
1998-2000
|
| Bowman Creek |
N/A
|
1
|
-
|
-
|
1998-2000
|
| Klahowya |
N/A
|
2
|
-
|
-
|
1998-2000
|
| Elbe Hills |
N/A
|
3
|
-
|
-
|
1999-2000
|
* radiotagged birds
† 1 = National Park; no history of commercial logging
2 = National Forest; low to moderate levels
of logging in the past
3 = State (Department of Natural Resources)
or private land; subjected to moderate to high levels of logging

Figure 1.1 Map of western Washington showing locations of 23 goshawk territories where the study took place. Asterisks denote weather stations; various circles represent the type(s) of data collected at each territory. At a minimum, reproductive output (o) was monitored at each territory.
![]()
|
(a)
|
 |
|
(b)
|
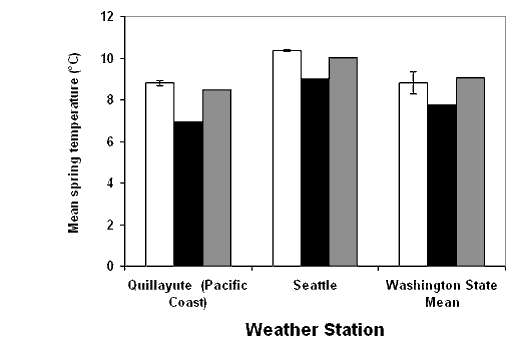 |
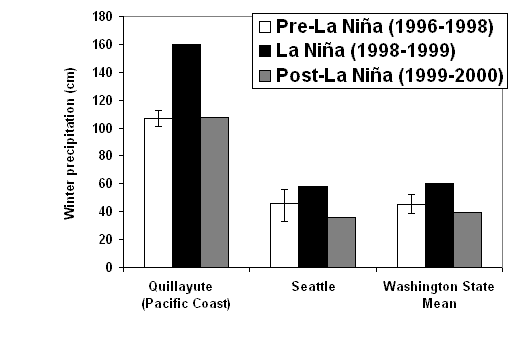
| Figure 1.2 Interannual variation in (a) winter precipitation and (b) spring temperature for two western Washington weather stations and the overall state mean. Bars indicate range of values during the pre-La Niña period of this study (National Climatic Data Center 1999a). | |
|
(a)
|
 |
|
(b)
|
 |
|
(c)
|
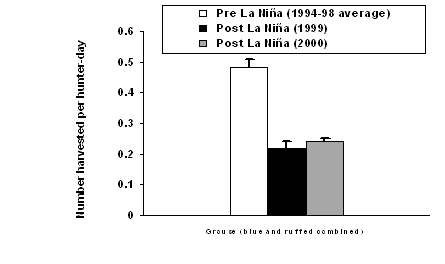 |
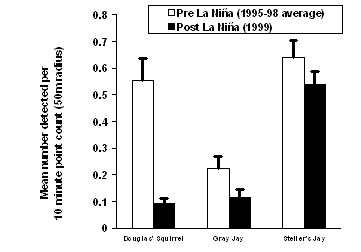
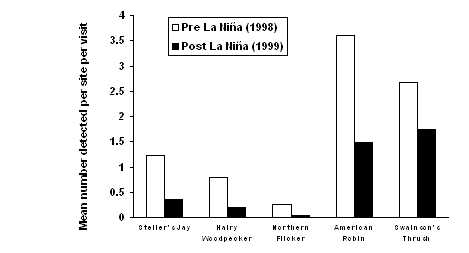
| Figure 1.3 Change in relative abundance of selected goshawk prey in western Washington following the La Niña winter of 1998-99. Data were obtained from three sources: (a) this study, (b) Bednarz et al. 2002, and (c) Washington Department of Fish and Wildlife 2000, 2001. Bars represent variation (SE) among (a) 27 study sites, or (c) 18 counties. | |
![]()
|
(a)
|
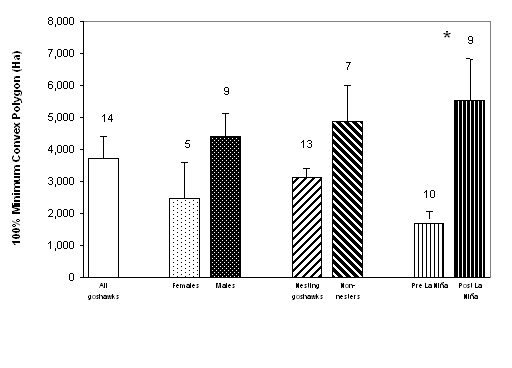 |
|
(b)
|
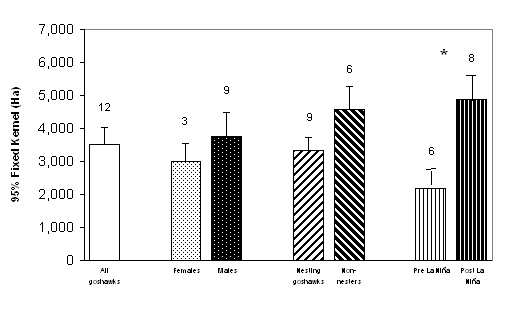 |
| Figure 1.4 Mean sizes of Northern Goshawk home ranges during the breeding season in managed forests of western Washington using (a) 100% minimum convex polygon (MCP) and, (b) 95% fixed kernel methods. Bars represent 1 SE (sample size shown above bars). Asterisk denotes significant difference (P < 0.05) between pairs. | |
|
(a)
|
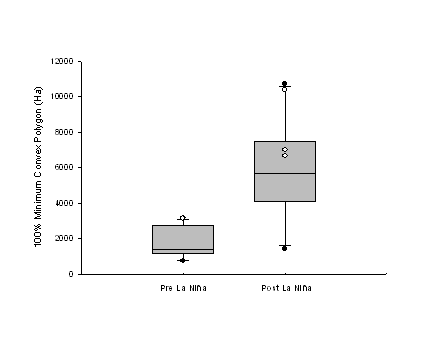 |
|
(b)
|
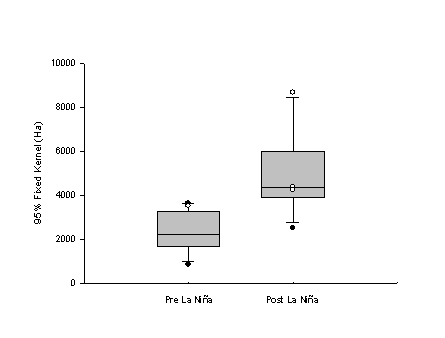 |
| Figure 1.5 Box and whisker plots showing differences between pre- and post-La Niña home range sizes of Northern Goshawks using the (a) 100% minimum convex polygon (MCP) and (b) 95% fixed kernel methods. White dots represent birds that either did not breed during one of the pre-La Niña years (n = 1) or did breed during one of the post-La Niña years (n = 3). Boxes depict the median score and 25% and 75% quartiles. Whiskers represent the 10th and 90th percentiles and black dots represent the 5th and 95th percentiles. | |
![]()
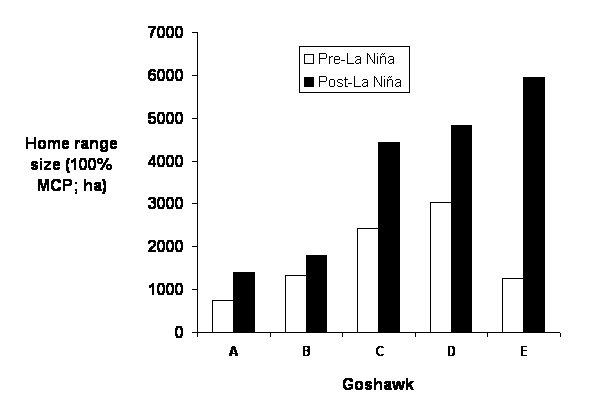 |
| Figure 1.6 Home range size before and after La Niña for five Northern Goshawks (A-E). |
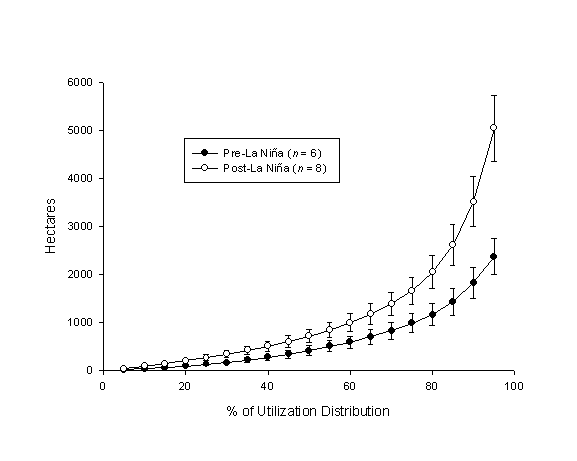 |
| Figure 1.7 Comparison of pre- and post-La Niña home ranges (95% fixed kernel) at 5% increments of the utilization distribution. |
![]()
|
(a)
|
 |
|
(b)
|
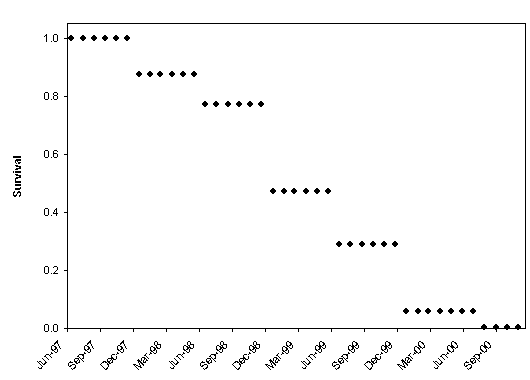 |
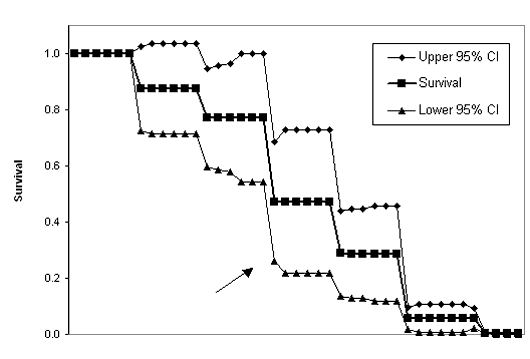
| Figure 1.8 Kaplan-Meier survival function (a) and pre-/post-La Niña fitted regression lines (ßpre = -0.017, ßpost = -0.029) of the survival function (b) for Northern Goshawks radiotagged in western Washington, June 1997-November 2000. The survival function was modified using staggered entry of animals. | |
 |
| Figure 1.9 Mean annual number of young fledged per occupied territory before and after La Niña for goshawk territories (n = 13) that were occupied in at least one pre-La Niña year and at least one post-La Niña year. Bars represent 1 SE. |
![]()
CHAPTER 2
Foraging Behavior and Habitat Use by Northern Goshawks in Managed
Forests of Western Washington
Abstract
I used radiotelemetry to study the foraging behavior and habitat
use patterns of Northern Goshawks (Accipiter gentilis) in
managed forests of western Washington from 1997-2000. Goshawks primarily
hunted using the 'short-stay perched-hunting' method, but also hunted
"on the wing" ('low soaring' method) when in areas dominated
by younger forest where below-canopy flight space was limited. Medium
to large birds such as Blue Grouse (Dendragapus obscurus),
Ruffed Grouse (Bonasa umbellus), and Band-tailed Pigeons
(Columba fasciata) dominated the diet of this goshawk population.
Important mammal species include snowshoe hare (Lepus americanus),
Douglas' squirrel (Tamiasciurus douglasii), and northern
flying squirrel (Glaucomys sabrinus). Of 104 identified prey
items, 75% were avian and 25% mammalian species. I identified 52
kill sites of 13 individual goshawks. Kills occurred in stands ranging
from relatively open, 13-yr-old regeneration units (pre-canopy stage)
to 200-yr-old complex, old-growth forests. Stands ranged from small
(0.5 ha) and isolated to large (> 100 ha) and contiguous. Goshawks
hunted in all forest types and successional stages except for recent
clearcuts and shrub/sapling stages. Univariate comparisons between
kill sites and paired random sites showed that goshawks tended to
hunt in stands with larger diameter trees and avoided areas composed
primarily of small trees (sapling/pole). Kill sites had an average
of 35% less tall (![]() 3 m) understory cover than random sites. Low (< 3 m) understory
cover varied considerably at both kill and random sites and did
not differ statistically. Managers of relatively young (< 60
yr) forests in western Washington can promote use of stands for
hunting by goshawks by thinning to reduce densities of small trees
and increase remaining tree diameters while maintaining canopy closure
and abundance of snags.
3 m) understory cover than random sites. Low (< 3 m) understory
cover varied considerably at both kill and random sites and did
not differ statistically. Managers of relatively young (< 60
yr) forests in western Washington can promote use of stands for
hunting by goshawks by thinning to reduce densities of small trees
and increase remaining tree diameters while maintaining canopy closure
and abundance of snags.
KEY WORDS: Accipiter gentilis, central place foraging, diet, foraging behavior, foraging habitat, forest structure, forest management, matched-pairs logistic regression, Northern Goshawk, thinning, Washington
In determining whether or not a population requires special conservation
attention, biologists generally either assess the state of the population
directly via demographic studies (by estimating ![]() )
or by assessing habitat use patterns to see if the population relies
on rare or declining habitats or habitat features within landscapes.
For species that are elusive and occur naturally at very low densities
it is often difficult to adequately quantify population size and
rate of change. Under these conditions, it is increasingly important
to conduct well-designed habitat studies throughout the population's
range, which may provide the necessary information to determine
what, if any, special measures need to be taken to conserve the
species. The Northern Goshawk (Accipiter gentilis) fits this
pattern. Goshawks have been at the center of debate between environmental
groups and biologists in western North America over the past decade.
At issue is whether or not goshawks require mature and old-growth
forests and whether or not logging has caused substantial declines
in the western U.S. goshawk population (Kennedy 1997).
)
or by assessing habitat use patterns to see if the population relies
on rare or declining habitats or habitat features within landscapes.
For species that are elusive and occur naturally at very low densities
it is often difficult to adequately quantify population size and
rate of change. Under these conditions, it is increasingly important
to conduct well-designed habitat studies throughout the population's
range, which may provide the necessary information to determine
what, if any, special measures need to be taken to conserve the
species. The Northern Goshawk (Accipiter gentilis) fits this
pattern. Goshawks have been at the center of debate between environmental
groups and biologists in western North America over the past decade.
At issue is whether or not goshawks require mature and old-growth
forests and whether or not logging has caused substantial declines
in the western U.S. goshawk population (Kennedy 1997).
While it is difficult to assess a population trend due to the nature
of goshawks (low density, difficult to survey over large areas,
etc.), biologists can conduct studies on habitat use of goshawks
to ascertain whether or not they rely on certain forest types and/or
successional stages for breeding, feeding, roosting, and dispersal.
In addition to identifying which habitats are used more or less
than expected based on occurrence, it is also important to study
how goshawks use various habitats (foraging behavior) so that we
understand the mechanisms that are determining their disproportionate
use (Marzluff et al. 2001).
Many studies have quantified habitats used by goshawks for nesting
in the western U.S. (see review in Squires and Reynolds 1997), but
few have addressed habitats used for foraging (Austin 1993, Bright-Smith
and Mannan 1994, Hargis et al. 1994, Beier and Drennan 1997, Good
1998). Further, no published studies have described the behavior
of goshawks while hunting in North America. Kenward (1978, 1982)
and Wide´n (1984, 1989) have written extensively on both topics
in Europe. Similarities between foraging habitat use in North America
and Europe include selection for older, closed-canopied forest containing
larger trees and greater basal area. Goshawk habitat use in Europe,
however, appears more varied with some populations adapted to anthropogenic
conditions such as edges of agriculture and woodland (Kenward 1982,
1996) and utilization of city landfills (Tornberg and Colpaert 2001).
To add to our understanding of goshawk ecology, I addressed the
question of foraging behavior and habitat use by Northern Goshawks
in the Pacific Northwest at the finest of scales by using radio-telemetry
to monitor hunting forays and to identify kill sites. My goal was
to determine whether or not goshawks require certain types of forest
conditions for hunting in managed forests of western Washington
and, if so, make specific recommendations to public and private
forest managers regarding goshawk habitat management. My objectives
were to: 1) describe the behavior of goshawks while hunting in managed
forests of western Washington, and 2) quantify foraging habitat
use in terms of within-stand forest structure. My null hypothesis
was that goshawks would hunt in all available forest conditions
equally, regardless of overstory and understory characteristics.
Finn et al. (2002a,b) suggest that understory development may impede
use of stands for nesting on the Olympic Peninsula of Washington.
Therefore, I was particularly interested in this variable with respect
to goshawk hunting.
![]()
METHODS
STUDY AREA
The study took place during the breeding period (April - August) from 1997-2000. I studied goshawks from nine territories throughout western Washington, primarily in the Olympic Mountains, Willapa Hills, and central Cascade Mountains west of the Cascade crest (Figure 2.1). Goshawks breed in this area from near sea level up through the mid elevations in the Cascade and Olympic ranges (nests in this study ranged from 120 - 750 m above sea level). A mild, moist, maritime climate supports dense conifer forests in the region with very high levels of forest productivity. Annual precipitation in the region is generally high on the windward sides of the Olympic and Cascade Mountains (250-365 cm); however, the northeast corner of the Olympic Peninsula is in a rain shadow and receives far less annual precipitation (80-150 cm; Franklin and Dyrness 1988). Douglas-fir (Pseudotsuga menziesii) and western hemlock (Tsuga heterophylla) are the most common overstory species in low elevation western Washington with western redcedar (Thuja plicata), Sitka spruce (Picea sitchensis), Pacific silver fir (Abies amabilis), and red alder (Alnus rubra) being common as well (Franklin and Dyrness 1988). Common understory and shrub species include western hemlock, western redcedar, swordfern (Polystichum munitum), salal (Gaultheria shallon), Oregon grape (Berberis nervosa), and red huckleberry (Vaccinium parvifolium). Within goshawk territories, natural openings (e.g. meadows, wetlands, rock outcrops) were rare and forest cover was relatively contiguous, albeit in varying stages of succession. The territories were on lands intensively managed for timber production (State or privately owned land), except for one that was on moderately managed land (National Forest). The intensively managed landscape was a mosaic of relatively discrete patches ranging from recent clearcuts to harvest age forest (i.e., 35-80 years). Old-growth forest was rare in all areas (< 1%) except in the one territory on National Forest where it covered an estimated 10-15% of the landscape.
RADIOTELEMETRY
Selection and capture of goshawks.
Forestry workers, falconers, or wildlife surveyors initially located
all of the territories used in this study (Finn 2000, Finn et al.
2002a,b). I returned to reported locations and used standard broadcast
methods (Kennedy and Stahlecker 1993) to locate active goshawk nests.
Adult goshawks were captured at their nests using Dho-gaza nets
and a live Great Horned Owl (Bubo virginianus) as lure (Bloom
et al. 1992). All hawks were fitted with an 18-g backpack style
transmitter-harness package (Buehler et al. 1995; 2-3% of body weight,
Advanced Telemetry Systems, Isanti, MN). Transmitter batteries lasted
up to three years and no premature failures were confirmed out of
35 birds radiotagged during the study. I returned to evaluate the
behavior of the bird within 72 hours to ensure that the transmitter
was not causing any obvious adverse effects (Withey et al. 2001).
Radiotracking. I generally
focused on a single goshawk for an entire day (0500-2100 hrs) to
document as many hunting locations as possible as well as to accurately
document its foraging behavior. Once a goshawk was located in the
morning I would home in on its location and stay with it (remaining
approximately 100-200m away) for as much of the day as possible,
identifying sites where kills were made, determining how long it
remained at each perch, how far it moved between perches, and other
attributes of foraging behavior. To confirm that a kill was made
I approached a hawk's location when the variation in the transmitter
pulse rate indicated it was feeding (mercury tipswitches caused
a slow pulse when vertical, fast when horizontal).
Determining kill locations.
While I was tracking, a video camera (with time stamp) was recording
the nest to confirm that prey was returned to the nest. During the
fledgling period, when young were in nest stands but not at nests,
I placed an observer in the stand to confirm prey deliveries. At
least half of the identified kill sites (55%) were confirmed directly
by either locating the prey remains or by seeing the goshawk leaving
the immediate area with prey in talons, but on the remainder of
occasions confirmation of the kill was not made until I reviewed
the nest video or communicated with the nest observer. In these
latter cases, the kill site location was estimated from my earlier
tracking locations. To calibrate these location estimates I conducted
a beacon test on transmitters hung in trees. The mean maximum error
estimate of these remaining kill sites was 69 m (range = 30-200).
I assumed that the goshawks last foraging position before a direct
flight back to the nest was the kill site unless I had contrary
evidence.
![]()
HABITAT ASSESSMENT
I collected stand structure data at all kill locations. I measured
overstory characteristics in two 0.04 ha circular plots. One plot
was centered on the site of the kill while the other was placed
100m away in the estimated direction of travel of the hawk before
making the kill. My goal was to sample the habitat the hawk flew
through on its way to making each kill. When I was not able to identify
the direction of travel (approximately 50% of the time) I placed
the second plot 100 m away in a random direction.
I measured the diameter at breast height (dbh) of all live conifers
and hardwoods ![]() 2.5
cm dbh. For tree density calculations I grouped trees into three
separate classes by dbh: Sapling/Pole: <30 cm; Medium Tree: 30-50
cm; Large Tree: >50 cm. I calculated basal area (m2/ha)
of all live conifers and hardwoods. I calculated the percent composition
of the overstory that was conifer versus hardwood by basal area.
I measured the dbh of all snags
2.5
cm dbh. For tree density calculations I grouped trees into three
separate classes by dbh: Sapling/Pole: <30 cm; Medium Tree: 30-50
cm; Large Tree: >50 cm. I calculated basal area (m2/ha)
of all live conifers and hardwoods. I calculated the percent composition
of the overstory that was conifer versus hardwood by basal area.
I measured the dbh of all snags ![]() 12.5
cm dbh and
12.5
cm dbh and ![]() 3
m in height. For snag density calculations I grouped snags into
two separate classes by dbh: Small Snags: 12.5-30 cm; Large Snags
>30 cm. I measured canopy closure (%) using a "Moosehorn"
vertical sighting tube (Robinson 1947) at each plot center as well
as 5 m out in each of the cardinal directions (10 measurements/kill
site).
3
m in height. For snag density calculations I grouped snags into
two separate classes by dbh: Small Snags: 12.5-30 cm; Large Snags
>30 cm. I measured canopy closure (%) using a "Moosehorn"
vertical sighting tube (Robinson 1947) at each plot center as well
as 5 m out in each of the cardinal directions (10 measurements/kill
site).
I measured understory vegetation along a 100 m line intercept transect
(Mueller-Dombois and Ellenberg 1974) extending between my two vegetation
plot centers. I recorded species, percent cover, and mean height
for every sapling (<12.5 cm dbh), shrub, and swordfern (Polystichum
munitum) that intercepted the line. I grouped understory plants
into low (<3 m) and tall (![]() 3 m) categories. I also measured percent cover of logs (
3 m) categories. I also measured percent cover of logs (![]() 30 cm diameter) and slash (<30 cm diameter wood) along understory
transects.
30 cm diameter) and slash (<30 cm diameter wood) along understory
transects.
Univariate Analysis
I randomly selected a site to pair with each kill location that
was 500-2,000 m from the site of the kill in a random direction.
This distance was chosen to insure that the paired site was in a
different stand, but within the same general part of the home range.
I compared paired sites that were in separate forest stands because
stands are relatively homogenous in managed forests. Also, I limited
selection of these sites to forested areas in which there were trees
![]() 12.5
cm dbh. This was done to exclude habitat types where I never detected
goshawks hunting (e.g. clearcuts, recently [<10 yr] planted stands,
water bodies). My goal was to compare stand structure at kill sites
to potentially available sites in the areas where the birds were
currently hunting.
12.5
cm dbh. This was done to exclude habitat types where I never detected
goshawks hunting (e.g. clearcuts, recently [<10 yr] planted stands,
water bodies). My goal was to compare stand structure at kill sites
to potentially available sites in the areas where the birds were
currently hunting.
I compared each stand structure variable between paired kill and
random sites using conditional logistic regression for 1:1 matched
data (Breslow and Day 1980, Hosmer and Lemeshow 1989, Hershey et
al. 1998). Hershey et al. (1998) provide a thorough description
of this method using Northern Spotted Owl (Strix occidentalis
caurina) nest sites to analyze stand structure variables similar
to mine.
Multivariate Analysis
I also used conditional logistic regression for 1:1 matched data to determine if there were particular combinations of variables that best distinguished kill sites from random sites (Hershey et al. 1998). To reduce the number of variables to be considered in a multivariate model, I excluded all variables with P values > 0.25 in univariate log-likelihood ratio tests. I further reduced the list of variables by analyzing correlations among them and retaining those that are more amenable to direct manipulation by forest managers. I also assessed interactions among variables in the multivariate analysis. I evaluated the full model of those remaining variables and only retained those variables whose P values again were < 0.25. I then used Akaikie's Information Criterion (AIC) to determine the most parsimonious model(s).
![]()
RESULTS
FORAGING BEHAVIOR
I observed two different hunting techniques and their relative
use appeared to be related to habitat type. 'Short-stay perched-hunting'
(Kenward 1982), where hawks occupy a perch, scan their surroundings
for prey, then fly to another perch (usually 100-300 m away), was
the most commonly used method overall (72% of foraging bouts) and
in stands > 30 yr old (small sawtimber stage and older; 96% of
foraging bouts). In addition to capturing adult and juvenile birds
and mammals using this method, I also witnessed two cases of nest
predation by goshawks (Steller's Jay [Cyanocitta stelleri]
and Varied Thrush [Ixoreus naevius]) as well as attacks on
fledglings of other raptor species at their nest sites (including
a successful kill of a Barred Owl [Strix varia]). In areas
dominated by younger, denser stages of sapling/pole forest (10-20
yr old), where below-canopy flight space is limited, goshawks tended
to use a 'low soaring' technique (13% of total foraging bouts).
This hunting technique was characterized by sustained flight <
100 m above the forest canopy generally in tight circles with little
flapping of the wings. I generally inferred this behavior by the
changing characteristics of the telemetry signal; however, on seven
occasions I witnessed the behavior (including attacks on prey).
In 15% of foraging bouts both types of hunting were utilized. This
generally occurred in highly fragmented areas where mature forest
and young, dense forest intermingle.
Medium to large birds such as Blue Grouse (Dendragapus obscurus),
Ruffed Grouse (Bonasa umbellus), and Band-tailed Pigeons
(Columba fasciata) dominated the diet of goshawks (Figure
2.2). Important mammal species include snowshoe hare (Lepus americanus),
Douglas' squirrel (Tamiasciurus douglasii), and northern
flying squirrel (Glaucomys sabrinus). Seventy-five percent
of prey items were avian and 25% were mammalian.
Breeding male goshawks returned to nests with prey every 4.8 ±
0.6 hr (n = 126 visits by nine birds) on average during daylight
hours. Non-breeding goshawks, however, generally consumed their
prey at the site of the kill over the course of a day or two. In
general, prey were captured and returned to nests immediately, except
in the case of larger prey items, which were always prepared extensively
before taken to the nest. Grouse (500-1,000 g) were decapitated,
plucked, and parceled into two pieces for separate deliveries, while
pigeons (360 g) were simply decapitated and plucked for a single
prey delivery. I never observed goshawks returning more than one
prey item at a time.
Males returned prey directly to nests, then females began feeding
young (during the early nestling stage). If females were not present
at nests, males waited there with prey until they returned (which
was never more than one minute in eight video observations). When
nestlings were large enough to feed themselves males dropped prey
at the nest and left immediately. Females were generally not on
the nest during this period, but would occasionally appear after
the young had fed and feed on the remaining portion, if available.
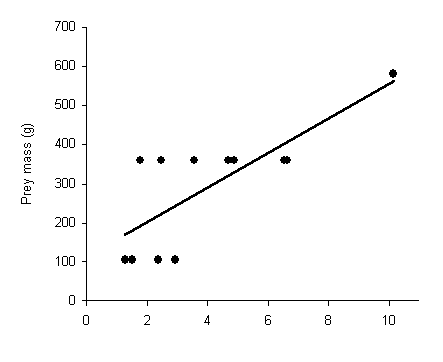
There was a positive relationship between size of prey returned
to the nest and the distance from nest where it was captured (n
= 12 cases for one male; r = 0.77, P = 0.003; Figure
2.3a). The gray shaded area in Figure 2.3b shows that if goshawks
traveled far from their nests (i.e. especially beyond about 4 km),
they did not return with small prey. In fact, as they traveled further
away larger and larger kills were required before returning to the
nest (n = 28 cases pooled across eight hawks; r =
0.42, P = 0.02; Figure 2.3b).
![]()
HABITAT
I identified 52 kill sites of 13 individual goshawks during the four-year study. Of these 13 hawks, seven were adult males, five were adult females, and one was a juvenile male. Kills occurred in stands ranging from relatively open, 13-yr-old regeneration units (pre-canopy stage) to 200-yr-old complex, old-growth forests. Stands ranged from small (0.5 ha) and isolated to large (> 100 ha) and contiguous. All forest types and successional stages were hunted except for recent clearcuts and shrub/sapling stages. Kills were made in later stages of forest development more than expected based on availability (x2 test of 'Young Forest' through 'Mature', df = 3, P < 0.001, Figure 2.4). There was substantial variation among kill sites in vegetative attributes (Table 2.1).
Univariate Analysis
In general, overstory variables were greater at kill sites - except
for small tree density and total tree density, which were greater
at random sites (Table 2.1). Goshawks tended to hunt in stands with
larger diameter trees and avoided areas composed primarily of small
trees (sapling/pole). Kill sites also had greater overall basal
area, greater total snag density and greater small snag density,
though large snag density did not differ between kill and random
sites. Canopy closure and percent conifer composition, which were
both very high throughout the study area, did not differ between
kill and random sites.
Most understory variables did not differ between kill and random
sites (Table 2.1). Understory cover only differed within the 'tall'
category (![]() 3 m; e.g. vine maple [Acer circinatum], conifer saplings,
salmonberry [Rubus spectabilis]). Kill sites had an average
of 35% less tall understory cover than random sites. Percent low
understory cover ranged widely at both kill and random sites, and
mean amounts of this habitat feature did not differ between sites.
Extreme sites were either dense, pole stage forests devoid of a
shrub layer, or mature stands with lush understories of swordfern
covering
3 m; e.g. vine maple [Acer circinatum], conifer saplings,
salmonberry [Rubus spectabilis]). Kill sites had an average
of 35% less tall understory cover than random sites. Percent low
understory cover ranged widely at both kill and random sites, and
mean amounts of this habitat feature did not differ between sites.
Extreme sites were either dense, pole stage forests devoid of a
shrub layer, or mature stands with lush understories of swordfern
covering ![]() 90%
of the forest floor. Understory species richness and percent cover
of both fine and coarse woody debris did not differ between kill
and random sites.
90%
of the forest floor. Understory species richness and percent cover
of both fine and coarse woody debris did not differ between kill
and random sites.
Multivariate Analysis
Many of the overstory variables were highly correlated with one
another as were some overstory with understory variables (e.g. small
tree density and tall understory cover). After eliminating insignificant
variables (P > 0.25 in univariate log likelihood ratio
tests) and further reducing the list of highly correlated variables,
eight remained for consideration (small tree density, medium tree
density, large tree density, mean dbh, basal area, canopy closure,
total snag density, and total understory cover). There were four
potential models that all had similarly low AIC values (Table 2.2).
Mean dbh was the only variable found in all four models - highlighting
its importance as a variable in predicting goshawk use of a stand
for hunting. Two other variables, basal area and total snag density,
also add predictive power to the model without substantially increasing
the AIC value. The model containing all three of these variables
had reasonable explanatory value with an R2 =
0.43.
I calculated the average ß for each of these variables over
the entire possible set of models using them (n = 7) based
on AIC weights (Burnham and Anderson 1998). Mean dbh was the most
important variable (ß = 0.104), followed by basal area (ß
= 0.012) and total snag density (ß = 0.004). The importance
of mean dbh is further highlighted when considering the odds ratios
for the 3-variable model (Table 2.3). The odds of selection for
a given stand increased by a factor of 2.3 as the mean dbh increased
from its mean at random sites (![]() random
= 24 cm) to its mean at kill sites (
random
= 24 cm) to its mean at kill sites (![]() kill
= 32 cm). Using the same model, the odds of selection increased
by a factor of 1.1 for both basal area (
kill
= 32 cm). Using the same model, the odds of selection increased
by a factor of 1.1 for both basal area (![]() random
= 40 m2/ha to
random
= 40 m2/ha to ![]() kill
= 52 m2/ha) and total snag density (
kill
= 52 m2/ha) and total snag density (![]() random
=45/ha to
random
=45/ha to ![]() kill
= 77/ha).
kill
= 77/ha).
![]()
DISCUSSION
FORAGING BEHAVIOR
The combination of hunting techniques utilized by this population
of goshawks is likely an adaptation to landscapes composed of a
diversity of habitat structures where a wide variety of prey is
available. The most commonly reported hunting style ('short-stay
perched-hunting' [Kenward 1982]) is generally used in the "typical"
goshawk foraging habitat (mature stands with large trees and sufficient
flight space beneath the canopy). In managed forests of western
Washington, however, where past and current forestry practices have
created a mix of relatively young habitat structural stages, adoption
of various hunting strategies allows goshawks to survive in landscapes
with very little "typical" habitat.
The overwhelming dominance of birds in this population's diet, particularly
larger birds such as grouse and Band-tailed Pigeons, is unusual
compared with other goshawk populations in North America (see review
in Squires and Reynolds 1997, Watson et al. 1998). Between 1986-96,
Watson et al. (1998) collected 531 prey items from 38 western Washington
goshawk nests. While grouse comprised a substantial portion of the
diet (16.5%), pigeons were completely absent from their prey collection.
Most of their nests were in National Forests or National Parks near
or above the Band-tailed Pigeons upper altitudinal limit (pigeons
are uncommon above ~ 1,000 m elevation in Washington). The nests
in my study (Figure 2.1) were mostly on State and private timberlands
at lower elevations (foothills) where this prey species is more
abundant.
The positive relationship I observed between prey size and distance
from nest is a prediction of Central Place Foraging (CPF) theory
(Orians and Pearson 1979) for 'single prey loaders' such as goshawks.
It appears that goshawks are non-selective with respect to prey
size within about 4 km of their nests, but beyond that distance
they require capture of relatively large prey before returning to
their nests (Figure 2.3). The 'benefits' of a given prey item must
outweigh the 'costs' of traveling such great distances and expenditure
of time for it to be returned to the nest. It is likely that hunting
goshawks did capture smaller prey when far away from nests, but
consumed them immediately, then continued foraging in the area until
a large prey item was captured.
HABITAT
In intensively managed forests of western Washington goshawks travel
great distances to secure prey in a wide variety of forested habitats,
particularly during periods of low prey abundance (Chapter 1). The
diversity of stand conditions utilized for hunting largely reflects
the diversity of habitats available within these managed landscapes.
While goshawks hunted throughout a range of stand structural conditions,
they used conditions representative of more mature forest development
more than expected based on availability. Young (<30 yrs) forests
generally do not provide the appropriate conditions (large trees
with well developed canopies, adequate flight space beneath the
canopy) for goshawk hunting.
This use of mature forests is similar to foraging studies on goshawks
elsewhere in the western United States. Beier and Drennan (1997)
found that goshawks in Arizona hunted in stands with greater densities
of large trees (>40.6 cm dbh) compared with contrast plots. Good
(1998) found that hunting goshawks in lodgepole pine (Pinus contorta)
forests of Wyoming returned more often to sites with greater densities
of large trees. In California, Hargis et al. (1994) detected goshawks
at sites with greater densities of large trees and greater basal
area relative to random points. Also in California, Austin (1993)
found goshawks selecting closed-canopied mature and old-growth stands.
The association I observed with snags may be explained by providing
additional perch sites while hunting an area using the 'short-stay
perched-hunting' method. In structurally simple managed forests
the presence of snags, even small ones, may provide strategic perch
locations to a hunting goshawk that would otherwise not be available.
Perhaps these small snags provide perches for hunting goshawks.
Large snags are rare in managed forests so my sampling methods were
probably not adequate to discern whether or not goshawks select
hunting areas based on these features.
Surprisingly, I did not find goshawk hunting limited to areas of
low to moderate shrub cover as expected. Finn et al. (2002a,b) found
higher occupancy rates at historic goshawk nests on the Olympic
Peninsula if shrub cover was relatively low in the stand (![]() = 19.0% at 12 occupied sites). However, I observed goshawks successfully
capturing prey in stands with up to 90% cover of a variety of shrub
species. It has been hypothesized (DeStefano and McCloskey 1997)
that the reason goshawks are rare in the Coast Ranges of Oregon
is due to dense understories making capture of prey difficult. My
results for low understory (i.e., shrubs <3 m high) do not support
this hypothesis for western Washington, where understories are very
similar to coastal Oregon. However, my finding of selection for
stands with lower than average tall understory (zone that encompasses
3m to generally about 6m high) does support it. It is likely that
goshawks require an open flight corridor beneath the canopy to be
successful in searching for, and capturing, prey more than they
need the ground layer to be free of shrubs. Tall understory was
highly correlated (r = 0.74) with small tree density so the
avoidance of areas with abundant tall understory may just be related
to goshawks selecting older stands (Figure 2.4), which have relatively
few small diameter trees. In recent years, xeric forests in eastern
Washington and throughout much of the Intermountain West have become
overstocked with small diameter trees in the understory due to nearly
a century of fire suppression (Agee 1993). This condition has likely
reduced the ability of goshawks to hunt in these forests, particularly
in younger stands where there is little flight space between the
overstory canopy and the shade tolerant understory conifers.
= 19.0% at 12 occupied sites). However, I observed goshawks successfully
capturing prey in stands with up to 90% cover of a variety of shrub
species. It has been hypothesized (DeStefano and McCloskey 1997)
that the reason goshawks are rare in the Coast Ranges of Oregon
is due to dense understories making capture of prey difficult. My
results for low understory (i.e., shrubs <3 m high) do not support
this hypothesis for western Washington, where understories are very
similar to coastal Oregon. However, my finding of selection for
stands with lower than average tall understory (zone that encompasses
3m to generally about 6m high) does support it. It is likely that
goshawks require an open flight corridor beneath the canopy to be
successful in searching for, and capturing, prey more than they
need the ground layer to be free of shrubs. Tall understory was
highly correlated (r = 0.74) with small tree density so the
avoidance of areas with abundant tall understory may just be related
to goshawks selecting older stands (Figure 2.4), which have relatively
few small diameter trees. In recent years, xeric forests in eastern
Washington and throughout much of the Intermountain West have become
overstocked with small diameter trees in the understory due to nearly
a century of fire suppression (Agee 1993). This condition has likely
reduced the ability of goshawks to hunt in these forests, particularly
in younger stands where there is little flight space between the
overstory canopy and the shade tolerant understory conifers.
![]()
Managing young stands for hunting goshawks
In this study area, goshawks used areas containing high densities
of small trees less than expected based on availability and used
areas composed of fewer, but larger, trees more than expected based
on availability. Additionally, kill sites had high canopy closures
(96% of kill sites had canopy closures ![]() 60%).
Goshawks may benefit if young stands in managed forests were thinned.
60%).
Goshawks may benefit if young stands in managed forests were thinned.
In research on goshawks in the same study area, Finn et al. (2002a,b)
recommend promoting nest stand development through moderate-level
thinning to 345 - 445 trees/ha across the range of diameter classes
in stands 30-35 years of age. At 5-10 years post-thinning, this
approach begins to yield the deep forest canopies and low shrub
cover found by Finn et al. (2002a,b) as important features of nest
stand habitat. Canopy cover would also increase over this time period.
Following the 5-10 year period of post-thinning response, treated
stands would serve as goshawk nest stand habitat for as long as
they were retained.
The thinning prescription recommended by Finn et al. for goshawk
nest stands would also suffice for creation of goshawk hunting habitat,
however, more emphasis should be on thinning "from below"
(removal of smallest trees first) to open up the understory without
sacrificing canopy closure too much. Flight space for goshawks would
improve immediately after harvest while canopy cover would improve
with time. Heavy thinning of 30-40 year old stands, which in western
Washington and Oregon is to tree densities of 148 - 247 trees/ha
(L. Raynes, pers. comm.), would likely reduce canopy closure to
levels too low to serve as goshawk habitat, even given the canopy
development that would occur post-thinning.
Employing variable density thinning (Carey and Johnson 1995, Carey
et al. 1999), as opposed to uniform prescriptions, would provide
a diversity of habitat niches supporting different prey species.
Leaving some small (< 1 ha) areas unthinned would provide the
dense, pole stages of forest that some key prey species thrive in
(e.g. snowshoe hare, ruffed grouse). While individual goshawks often
specialize on one or a few prey species (Figure 2.3a), a diverse
prey assemblage may be important in maintaining goshawk populations
during times of environmental stress (Chapter 1).
Commercial thinning generally reduces small snag density over the
development pathway of a stand by removing trees that would otherwise
die due to competitive mortality. Also, during logging snags can
be inadvertently damaged or actively removed due to safety or logistical
considerations (Hayes et al. 1997). These losses may be ameliorated
by the effect that thinning has on the rate of snag creation from
wind in stands. Because neighboring trees are removed and can no
longer serve as wind buffers, trees in commercially thinned stands
are more susceptible to damage to their tops and blowdown (J. Clarke,
pers. comm.). Even so, it may be important to create snags in landscapes
that are subject to widespread commercial thinning. Increasing snag
density in young, managed forests could be achieved during commercial
thinning operations through various methods (e.g., girdling, fungal
inoculation, topping with mechanical harvesters; Lewis 1998).
Finally, allowing a portion of the managed forest landscape to mature
beyond industrial rotation ages (currently 40-50 years on the Olympic
Peninsula, L. Raynes, pers. comm.) would ensure that a wide range
of goshawk hunting habitats are available within a particular area.
On the Olympic Peninsula, extending rotations would be particularly
useful and economically practicable (because fewer acres would be
necessary) in areas near late successional reserves of Olympic National
Park or Olympic National Forest where extant habitat can be augmented
and more late successional habitat is planned (Finn et al. 2002a,
USDA, USDI 1994).
![]()
Table 2.1. Vegetative attributes at 52 goshawk kill sites and 52 randomly located sites within territories in managed forests of western Washington.
|
Variable
|
Kill sites
|
Random
|
|||||
|
Mean
|
SE
|
Range
|
Mean
|
SE
|
Range
|
P <
|
|
| Overstory | |||||||
| Total tree
density (#/ha; all live stems |
800
|
84
|
235-4115
|
1308
|
175
|
248-6369
|
0.009
|
| Small tree density (#/ha; <30 cm dbh) |
514
|
90
|
50-4115
|
1107
|
185
|
54-6338
|
0.002
|
| Medium tree density (#/ha; 30-50 cm dbh) |
225
|
19
|
0-495
|
160
|
17
|
0-408
|
0.005
|
| Large tree density (#/ha; >50 cm dbh) |
62
|
7
|
0-235
|
41
|
7
|
0-223
|
0.035
|
| Mean dbh (cm) |
32
|
1
|
Oct-56
|
24
|
1
|
Apr-42
|
0.001
|
| Basal area (m2/ha) |
52
|
2
|
Sep-90
|
40
|
2
|
Jun-70
|
0.001
|
| Canopy closure (%) |
77
|
1
|
35-90
|
75
|
2
|
36-91
|
0.206
|
| Total snag
density (#/ha; all snags |
77
|
10
|
0-285
|
45
|
10
|
0-384
|
0.022
|
| Small snag density (#/ha; 12.5-30 cm dbh) |
63
|
9
|
0-285
|
36
|
9
|
0-371
|
0.035
|
| Large snag density (#/ha; >30 cm dbh) |
14
|
3
|
0-74
|
9
|
3
|
0-124
|
0.201
|
| Percent conifer (by basal area) |
94
|
2
|
46-100
|
92
|
2
|
15-100
|
0.5
|
| Understory | |||||||
| Total understory cover (%) |
36
|
4
|
Feb-90
|
45
|
4
|
0-97
|
0.109
|
| Low understory cover (%; <3m) |
25
|
3
|
0-90
|
26
|
3
|
0-91
|
0.887
|
| Tall understory
cover (%; |
15
|
2
|
0-68
|
23
|
4
|
0-85
|
0.047
|
| Understory species richness (spp./100 m line transect) |
5
|
1
|
20-Jan
|
6
|
1
|
0-16
|
0.27
|
| Slash cover (%; woody debris <30 cm diameter) |
11
|
2
|
0-75
|
10
|
2
|
0-45
|
0.528
|
| Log cover
(%; woody debris |
4
|
1
|
0-9
|
3
|
1
|
0-17
|
0.513
|
Table 2.2. Akaike's Information Criterion (AIC) values for four competing goshawk foraging habitat models.
|
Model
|
AIC
|
| Mean dbh |
55.5
|
| Mean dbh + Snag density |
55.9
|
| Mean dbh + Basal area |
57
|
| Mean dbh + Basal area + Snag density |
57.8
|
|
ß
|
Odds ratios
|
|
| Single variable model 1 | ||
| Mean dbh |
0.11
|
2.5
|
| Full model (3 variables) 2 | ||
| Mean dbh |
0.098
|
2.3
|
| Basal area |
0.008
|
1.1
|
| Snag density |
0.004
|
1.1
|
| 1
R2 = 0.40 2 R2 = 0.43 |
![]()
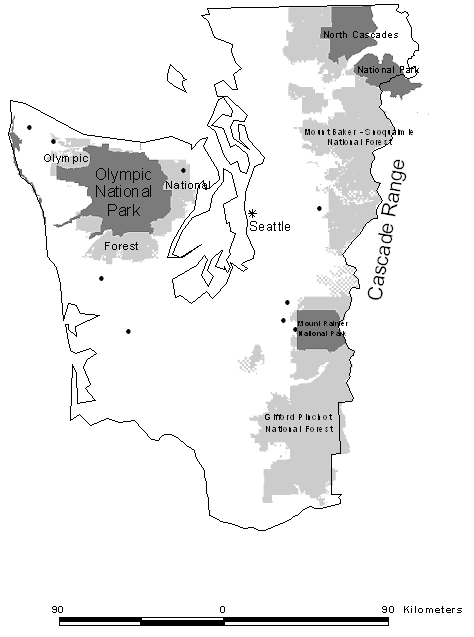 |
| Figure 2.1 Map of western Washington showing locations of nine goshawk territories (black dots) where foraging study took place in relation to major land ownership patterns (non-shaded areas are predominately private and state forestlands). |
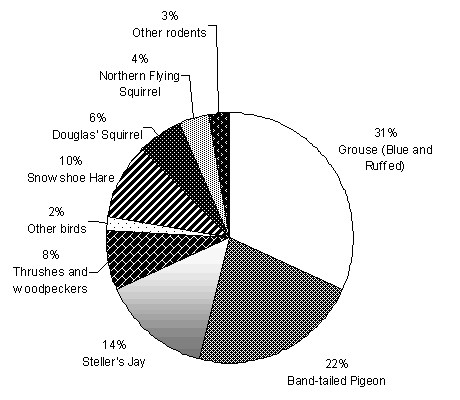 |
| Figure 2.2. Diet of goshawks in western Washington from 1997-2000 (n = 104 prey items from 15 territories). Prey items were identified by collecting remains (fur, feathers, and bones) in nest stands or at kill sites, or by reviewing video footage from nest cameras. |
![]()
|
(a)
|
 |
|
(b)
|
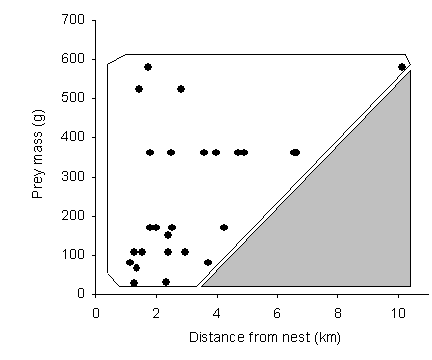 |
| Figure 2.3. Relationship between size of prey and the distance away from the nest where it was captured for (a) a single male with 12 kills, and (b) all goshawks combined (n = 28 cases pooled across eight hawks). Only those prey that were returned to the nest are included. | |
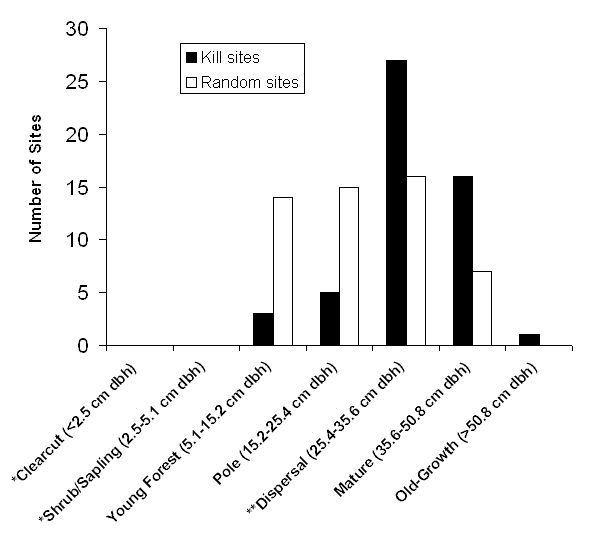 |
| Figure 2.4 Forest structural stages at 52 goshawk kill sites and 52 randomly located sites within territories in managed forests of western Washington, 1997-2000. Structural stages are adapted from Oliver et al. (1995). *All 'Clearcut' and 'Shrub/Sapling' stage stands were excluded from the random sample because hunting goshawks did not use these stages of forest development. ** 'Dispersal' forest refers to the minimum stage required by Northern Spotted Owls for foraging and dispersal with protection from predators. Size ranges are in quadratic mean diameter (QMD). |
![]()
LITERATURE CITED
Agee, J. K. 1993. Fire ecology of Pacific Northwest forests.
Island Press, Washington,
D.C.
Austin, K. K. 1993. Habitat use and home range size of breeding
Northern Goshawks in
the southern Cascades. M.Sc. thesis, Oregon State University,
Corvallis, OR.
Bayer, R. D. 1986. Breeding success of seabirds along the mid-Oregon
coast concurrent
with the 1983 El Niño. Murrelet 67:23-26.
Bednarz, J. C., D. M. Juliano, M. J. Huss, and D. E. Varland.
2002. Interactions between
fungi and cavity nesters: Applications to managing wildlife
and forests in western
Washington: 1999 Annual Report. Dept. Biol. Sci., Arkansas State
Univ., State
University, AR.
Beier, P., and J. E. Drennan. 1997. Forest structure and prey
abundance in foraging areas
of northern goshawks. Ecological Applications 7:564-571.
Bloom, P. H., E. H. Henckel, J. K. Schmutz, B. Woodbridge, J.
R. Bryan, R. L. Anderson,
P. J. Detrich, T. L. Maechtle, J. O. McKinley, M. D. McCrary,
K. Titus, and P. F.
Schempf. 1992. The dho-gaza with Great Horned Owl lure: an analysis
of its
effectiveness in capturing raptors. Journal of Raptor Research
26:167-178.
Boal, C. W., D. E. Andersen, and P. L. Kennedy. 2000. Breeding
season home range and
habitat use of male Northern Goshawks (Accipiter gentilis)
in Minnesota. Papers
presented at the Raptor Research Foundation Annual Meeting,
Jonesboro, AR,
November 2000.
Boersma, P. D. 1998. Population trends of the Galapagos Penguin:
Impacts of El Niño and
La Niña. Condor 100:245-253.
Breslow, N. E., and N. E. Day. 1980. Statistical methods in
cancer research. Volume 1. The
analysis of case-control studies. International Agency for Research
on Cancer,
Scientific Publication 32, Lyon, France.
Bright-Smith, D. J., and R. W. Mannan. 1994. Habitat use by
breeding male Northern
Goshawks in northern Arizona. Studies in Avian Biology 16:58-65.
Buehler, D. A., J. D. Fraser, M. R. Fuller, L. S. McAllister,
and J. K. D. Seegar. 1995.
Captive and field tested radio attachment techniques for Bald
Eagles. Journal of
Field Ornithology 66:173-180.
![]()
Burnham, K. P., and D. R. Anderson. 1998. Model selection and
inference: a practical
information-theoretic approach. Springer, New York, NY.
Cane, M. A. 1983. Oceanographic events during El Niño.
Science 222:1189-1195.
Carey, A. B., and M. L. Johnson. 1995. Small mammals in managed,
naturally young, and
old-growth forests. Ecological Applications 5:336-352.
Carey, A. B., J. Kershner, B. Biswell, and L. D. de Toledo.
1999. Ecological scale and
forest development: squirrels, dietary fungi, and vascular plants
in managed and
unmanaged forests. Wildlife Monographs 142:1-71.
DeStefano, S., and J. McCloskey. 1997. Does vegetation structure
limit the distribution of
northern goshawks in the Oregon Coast Ranges? Journal of Raptor
Research
31:34-39.
Elkins, N. 1983. Weather and bird behaviour. T & A.D. Poyser
Ltd., Staffordshire,
England.
Finn, S. P. 2000. Multi-scale habitat influences on Northern
Goshawk occupancy and
reproduction on Washington's Olympic Peninsula. M.Sc. thesis,
Boise State
University, Boise, ID.
Finn, S. P., J. M. Marzluff, and D. E. Varland. 2002a. Effects
of landscape and local
habitat attributes on Northern Goshawk site occupancy in western
Washington.
Forest Science 48:427-436.
Finn, S. P., D. E. Varland, and J. M. Marzluff. 2002b. Does
Northern Goshawk nest
stand occupancy vary with nest stand characteristics on the
Olympic Peninsula,
Washington? Journal of Raptor Research 36:265-279.
Franklin, J. F., and C. T. Dyrness. 1988. Natural vegetation
of Oregon and Washington.
Oregon State University Press, Corvallis, OR.
Good, R. E. 1998. Factors affecting the relative use of northern
goshawk (Accipiter
gentilis) kill areas in southcentral Wyoming. M.Sc. thesis,
University of Wyoming,
Laramie, WY.
Grant, P. R., B. R. Grant, L. F. Keller, and K. Petren. 2000.
Effects of El Niño events
on Darwin's Finch productivity. Ecology 81:2442-2457.
Gremel, S. 2001. Spotted Owl monitoring in Olympic National
Park: 2001 annual report.
U.S. Dept. Int., National Park Service, Olympic National Park,
Port Angeles, WA.
Guerra, C. G., L. C. Fitzpatrick, R. Aguilar, and B. J. Venables.
1988. Reproductive
consequences of El Niño - southern oscillation in gray
gulls (Larus modestus).
Colonial Waterbirds 11:170-175.
Hargis, C. D., C. McCarthy, and R. D. Perloff. 1994. Home ranges
and habitats of
Northern Goshawks in eastern California. Studies in Avian Biology
16:66-74.
Hatch, S. A. 1987. Did the 1982-1983 El Niño-Southern
Oscillation affect seabirds in
Alaska? Wilson Bulletin 99:468-474.
![]()
Hayes, J. P., S. S. Chan, W. H. Emmingham, J. C. Tappeiner,
L. D. Kellogg, and
J. D. Bailey. 1997. Wildlife response to thinning young forests
in the Pacific
Northwest. Journal of Forestry 95:28-33.
Hershey, K. T., E. C. Meslow, and F. L. Ramsey. 1998. Characteristics
of forests at
spotted owl nest sites in the Pacific Northwest. Journal of
Wildlife Management
62:1398-1410.
Hodder, J., and M. R. Graybill. 1985. Reproduction and survival
of seabirds in Oregon
during the 1982-1983 El Niño. Condor 87:535-541.
Hooge, P. N., and B. Eichenlaub. 1997. Animal movement extension
to ArcView: version
1.1. Alaska Biol. Sci. Ctr., U.S. Geol. Survey, Anchorage, AK.
Hosmer, D. W., and S. Lemeshow. 1989. Applied logistic regression.
John Wiley, New
York, NY.
Ingraldi, M. F. 1998. Population biology of Northern Goshawks
in east-central Arizona.
Nongame and Endangered Wildlife Program Technical Report 133.
Arizona Game
and Fish Department, Phoenix, AZ.
Iverson, G. C., G. D. Hayward, K. Titus, E. DeGayner, R. E.
Lowell, D. C.
Crocker-Bedford , P. F. Schempf, and J. Lindell. 1996. Conservation
assessment
for the Northern Goshawk in southeast Alaska. Gen. Tech. Rep.
PNW-GTR-387,
U.S. Dept. Agric., Forest Service, Pacific Northwest Research
Station, Portland,
OR.
Jaksic, F. M., S. I. Silva, P. L. Meserve, and J. R. Gutierrez.
1997. A long-term study of
vertebrate predator responses to an El Niño (ENSO) disturbance
in western South
America. Oikos 78:341-354.
Keane, J. J. 1999. Ecology of the Northern Goshawk in the Sierra
Nevada, California.
PhD dissertation, University of California, Davis, CA.
Kennedy, P. L. 1997. The northern goshawk (Accipiter gentilis
atricapillus): is there
evidence of a population decline? Journal of Raptor Research
31:95-106.
Kennedy, P. L., and D. W. Stahlecker. 1993. Responsiveness of
nesting Northern
Goshawks to taped broadcasts of 3 conspecific calls. Journal
of Wildlife
Management 57:249-257.
Kennedy, P. L., J. M. Ward, G. A. Rinker, and J. A. Gessaman.
1994. Post-fledging
areas in Northern Goshawk home ranges. Studies in Avian Biology
16:75-82.
Kenward, R. E. 1978. Hawks and doves: factors affecting success
and selection in
goshawk attacks on woodpigeons. Journal of Animal Ecology 47:449-460.
Kenward, R. E. 1982. Goshawk hunting behaviour, and range size
as a function of
food and habitat availability. Journal of Animal Ecology 51:69-80.
Kenward, R. E. 1996. Goshawk adaptation to deforestation: Does
Europe differ
from North America? Pages 233-243 in D. M. Bird, D. E. Varland,
and
J. J. Negro [Eds.], Raptors in human landscapes: Adaptations
to built and
cultivated environments. Academic Press, San Diego, CA.
Kenward, R.E., V. Marcstrom, and M. Karlbom. 1993. Causes of
death in
radio-tagged Northern Goshawks. Pages 57-61 in P. Redig, J.
Cooper, D.
Remple, D. Hunter and T. Hahn [Eds.], Raptor biomedicine. University
of
Minnesota Press, Minneapolis, MN.
Kernohan, B. J., R. A. Gitzen, and J. J. Millspaugh. 2001. Analysis
of animal space
use and movements. Pages 126 - 166 in J. J. Millspaugh and J.
M. Marzluff,
editors. Radio tracking and animal populations. Academic Press,
San Diego, CA.
Lewis, J. C. 1998. Creating snags and wildlife trees in commercial
forest landscapes.
Western Journal of Applied Forestry 13:97-101.
![]()
Luginbuhl, J. M., J. M. Marzluff, J. E. Bradley, M. G. Raphael,
and D.E. Varland. 2001.
Corvid survey techniques and the relationship between corvid
relative abundance
and nest predation. Journal of Field Ornithology 72:556-572.
Marzluff, J. M., S. T. Knick, and J. J. Millspaugh. 2001. High-tech
behavioral ecology:
modeling the distribution of animal activities to better understand
wildlife space use
and resource selection. Pages 310 - 326 in J. J. Millspaugh
and J. M. Marzluff,
editors. Radio tracking and animal populations. Academic Press,
San Diego, CA.
Marzluff, J. M., S. T. Knick, M. S. Vekasy, L. S. Schueck, and
T. J. Zarriello. 1997.
Spatial use and habitat selection of golden eagles in southwestern
Idaho.
Auk 114:673-687.
Massey, B. W., D. W. Bradley, and J. L. Atwood. 1992. Demography
of a California
Least Tern colony including effects of the 1982-1983 El Niño.
Condor 94:976-983.
McClaren, E. L., P. L. Kennedy, and S. R. Dewey. 2002. Do some
northern goshawk
nest areas consistently fledge more young than others? Condor
104:343-352.
Miskelly, C. M. 1990. Effects of the 1982-83 El Niño
event on two endemic landbirds
on the Snares Islands, New Zealand. Emu 90:24-27.
Mohr, C. O. 1947. Table of equivalent populations of North American
small mammals.
American Midland Naturalist 37:223-249.
Mueller-Dombois, D., and H. Ellenberg. 1974. Aims and methods
of vegetation ecology.
John Wiley, New York, NY.
National Centers for Environmental Prediction/Climate Prediction
Center. 1999. Cold and|
warm episodes by season. <http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.html>
National
Climatic Data Center. 1999a. U.S. climate at a glance.
(http://lwf.ncdc.noaa.gov/oa/climate/research/cag3/cag3.html)
National Climatic Data Center. 1999b. December 1998-February
1999: The Climate
in Historical Perspective.
http://www.ncdc.noaa.gov/ol/climate/research/1999/win/win99.html
Oliver, C., R. Greggs, L. Hicks, and S. Boyd. 1995. Forest stand
structural classification
system developed for the Plum Creek Cascades Habitat Conservation
Plan.
Technical Report No. 10, Plum Creek Timber Company, L.P., Seattle,
WA.
Orians, G. H., and N. E. Pearson. 1979. On the theory of central
place foraging.
Pp. 155-177 In D. J. Horn, G. R. Stairs, and R. D. Mitchell
(eds.), Analysis of
ecological systems. Ohio State University Press, Columbus, OH.
Otis, D. L., and G. C. White. 1999. Autocorrelation of location
estimates and the analysis
of radiotracking data. Journal of Wildlife Management 63:1039-1044.
Pollock, K. H., S. R. Winterstein, C. M. Bunck, and P. D. Curtis.
1989. Survival analysis
in telemetry studies: the staggered entry design. Journal of
Wildlife Management
53:7-15.
Reynolds, R. T., and S. M. Joy. 1998. Distribution, territory
occupancy, dispersal, and
demography of Northern Goshawks on the Kaibab Plateau, Arizona.
Final
Report for Arizona Game and Fish Heritage Project No. I94045,
U.S. Dept.
Agric., Forest Service, Rocky Mountain Research Station, Ft.
Collins, CO.
Robinson, M. W. 1947. An instrument to measure forest crown
cover. Forest Chronicle
23:222-225.
![]()
Schueck, L. S., and J. M. Marzluff. 1995. Influence of weather
on conclusions about
effects of human activities on raptors. Journal of Wildlife
Management 59:674-682.
Seaman, D. E., J. J. Millspaugh, B. J. Kernohan, G. C. Brundige,
K. J. Raedeke,
and R. A. Gitzen. 1999. Effects of sample size on kernel home
range
estimates. Journal of Wildlife Management 63:739-747.
Sillett, T. S., R. T. Holmes, and T. W. Sherry. 2000. Impacts
of a global climate
cycle on population dynamics of a migratory songbird. Science
288:2040-2042.
Squires, J. R., and R. T. Reynolds. 1997. Northern Goshawk (Accipiter
gentilis).
In The birds of North America, No. 298, Poole, A., and F. Gill,
[eds.].
The Academy of Natural Sciences, Philadelphia, PA, and The American
Ornithologists' Union, Washington, D.C.
Tornberg, R., and A. Colpaert. 2001. Survival, ranging, habitat
choice and diet of
the Northern Goshawk Accipiter gentilis during winter in Northern
Finland.
Ibis 143:41-50.
USDA, Forest Service; USDI, Bureau of Land Management. 1994.
Record of
decision for amendments to Forest Service and Bureau of Land
Management
planning documents within the range of the Northern Spotted
Owl. Standards
and guidelines for management of habitat for late-successional
and old-growth
forest related species within the range of the Northern Spotted
Owl.
Washington, D.C.
Vulgares, S., N. Vulgares, and J. P. Smith. 2000. Goshawk nesting
surveys in the
Olympic National Forest: May-August 1999. Report submitted to
U.S. Dept.
Agric., Forest Service, Olympic National Forest. Hawkwatch International,
Salt Lake City, UT.
Washington Department of Fish and Wildlife. 2000. 1999 Game
harvest report.
Wildl. Manage. Prog., Wash. Dept. Fish and Wildl., Olympia,
WA.
Washington Department of Fish and Wildlife. 2001. 2000 Game
harvest report.
Wildl. Manage. Prog., Wash. Dept. Fish and Wildl., Olympia,
WA.
Watson, J. W., D. W. Hays, S. P. Finn, and P. Meehan-Martin.
1998. Prey of
breeding Northern Goshawks in Washington. Journal of Raptor
Research
32:297-305.
White, G. C., and R. A. Garrott. 1990. Analysis of wildlife
radio-tracking data.
Academic Press, San Diego, CA.
Wide´n, P. 1984. Activity patterns and time budget in
the goshawk Accipiter gentilis
in a boreal forest area in Sweden. Ornis Fennica 61:109-112.
Wide´n, P. 1989. The hunting habitats of goshawks Accipiter
gentilis in boreal
forests of central Sweden. Ibis 131:205-213.
Wilson, U. W. 1991. Responses of three seabird species to El
Niño events and
other warm episodes on the Washington coast, 1979-1990. Condor
93:853-858.
Withey, J. C., T. D. Bloxton, and J. M. Marzluff. 2001. Effects
of tagging and
location error in wildlife radiotelemetry studies. Pages 43
- 75 in J. J.
Millspaugh and J. M. Marzluff, [eds.], Radio tracking and animal
populations.
Academic Press, San Diego, CA.
Worton, B. J. 1989. Kernel methods for estimating the utilization
distribution in
home-range studies. Ecology 70:164-168.
Younk, J. V., and M. J. Bechard. 1994. Breeding ecology of the
Northern
Goshawk in high-elevation aspen forests of northern Nevada.
Studies
in Avian Biology 16:119-121.
Zar, J. H. 1996. Biostatistical analysis: Third edition. Prentice-Hall,
Upper Saddle
River, N.J.
![]()

